THE BRETTANOMYCES II. TAXONOMIC SIGNIFICANCE OF SLOW FERMENTATIONS AND DESCRIPTION OF A NEW SPECIES
by R. B. Gilliland, B.A., B.Sc, F.R.I.C.
{Arthur Guinness Son & Co. [Dublin], Ltd., St. James’s Gate, Dublin}
Received 16th August, 1961
A number of strains of Brettanomyces, isolated from spoiled beer, were similar to one another but differed significantly from the hitherto described species. The characteristics of these strains warranted their separation into a new species for which the name Brettanomyces dublinensis is proposed. This yeast fermented some sugars very slowly and other examples of very slow fermentation were found in the genus Brettanomyces. Methods for the detection of slow or delayed fermentations have been compared and their taxonomic significance has been considered.
Introduction
In a previous paper3 the isolation of 44 strains of Brettanomyces from trade samples of beer was described. Of these strains, 32 corresponded to one or other of the established species Brett. bruxellensis or Brett. sckanderlii, but 11 strains, similar to one another in biochemical characteristics, differed appreciably from any established species. When the tests described by Lodder & Kreger-van Rij8 for the characterization of yeast species were applied to a typical representative of these strains, it was found that the new strains were most similar to Brett. anomalus but that they differed from this species in their ability to ferment sucrose. A detailed comparison with an authentic strain of Brett. anomalus, under more exacting conditions, revealed a number of slow or delayed fermentations. As the taxonomic significance of such fermentations is in dispute, this problem will be discussed before the results are considered.
The ability of a yeast to ferment specific carbohydrates is of primary importance in its classification. In most cases the use of either Einhorn or Durham tubes gives an unambiguous + or – result, but in some cases a very slow or delayed fermentation may be observed. Confusion is often caused by such fermentations as there is no unanimity of opinion, either on the precise form of the test, or on the taxonomic value of these slow fermentations. These problems, which have recently been discussed by Roberts & van der Walt,12 and by Pappagianis and Phaff,11 are of such importance that it is worth considering them in detail.
In the standard work on yeast classification, Lodder & Kreger-van Rij8 recommended the use of Einhorn tubes for the determination of fermentative ability. These tubes were incubated for 10 days at 25° C. Later the same authors9 advised the use of Durham tubes and these are now used by most workers; thus, Wickerham17 advised the use of Durham tubes with incubation at 25° C. for 24 days. The Durham tube and Einhorn tube methods are not equivalent, and neither may agree with quantitative tests using the van Iterson-Kluyver fermentometer, the Warburg fermentometer, or with tests based on determination of the amount of fermentation by chemical or physical methods; e.g., Stelling-Dekker14 found that the yeast Zygosaccharomyces japonicus (syn. Saccharomyces rouxii) was unable to ferment sucrose as judged by Einhorn tubes, but that it gave a slow non-delayed fermentation in the van Iterson-Kluyver apparatus, whereas Pappagianis & Phaff11 found that this yeast gave fast gas production after a long delay in Durham tubes. Thus, we can have negative, slow non-delayed, or fast delayed results for this yeast using different methods of examination.
It is sometimes not realized that Einhorn or Durham tubes may fail to detect a slow fermentation. If gas is to appear in the closed end or in the insert tube, it must be produced at a rate faster than it can diffuse from the open surface of the liquid. It is quite possible with slow fermentations to observe a small bubble in the inverted tube one day and for this bubble to disappear by the next day, and this type of fermentation can readily be missed altogether. A reliable quantitative method of determining whether or not fermentation has taken place would be of value in such cases.
The mechanism of slow fermentations has been considered by Pappagianis & Phaff,11 who studied the slow sucrose fermentations given by Zygosacch. japonicus and similar yeasts in some detail. They found that gas first appeared in Durham tubes after 20-30 days, and that the insert tube filled with gas during the few days after its first appearance. A subculture from an actively fermenting tube into a fresh tube of sucrose again required more than 20 days before gas was produced. In the van Iterson-Kluyver apparatus a heavy inoculum gave a slow but not delayed fermentation. No mutant cells could be detected in a culture of the yeast when it was plated on sucrose agar. Dried or frozen cells gave active inversion of sucrose in contrast to fresh cells, and aged cells fermented sucrose with less delay than fresh cells. These facts led the authors to deduce that the delayed fermentation was due to a change in cell wall permeability which took place on ageing or on drying; it was not considered to be due to mutation or induced enzyme formation.
A similar explanation may apply to the slow maltose-fermenters isolated as single spore cultures from a hybrid yeast by Gilliland.4 One strain which, when isolated, required 21 days to show gas production from maltose in Durham tubes, still required 15 days for gas production after 20 subcultures in maltose medium. When the fermentation of maltose by this strain was followed by daily determination of the specific gravity of the medium it was found that fermentation was very slow but quite appreciable during the first 20 days; the rate of fermentation then increased, and it was not until this stage that gas production was observed. This slow fermentation could not be due to adaptation, or to mutation, but is a stable property of the yeast. It should be noted that what appeared to be delayed fermentation in Durham tubes was again shown to be in reality a slow fermentation, followed by a rapid fermentation at the time of gas evolution.
Roberts & van der Walt12 on the otherhand found that with Sacch. capensis an apparently slow maltose fermentation was due to mutation of a gene m → M. In this case if the culture was pre-grown on a maltose medium it gave rapid fermentation of maltose. It was demonstrated that two distinct types of cell were present, one a maltose fermenter and one a non-fermenter. When a hybrid was formed between these two types the spore progeny of this hybrid yielded 2:2 segregation of maltose-fermenting ability. Similar results were found by Kossikov8 in the case of delayed fermentations of maltose and sucrose by Sacch. globosus and delayed maltose fermentations by Sacch. paradoxus. In each case fermentation occurred after a very long delay in the specified sugar, but a second subculture into the same sugar medium gave rapid fermentation of this sugar. Genetic analysis showed that there was mutation of a gene from the recessive to the dominant form, and the author argued that this mutation was directed by the specific substrate.
Takahashi & Ikeda15 found slow and very slow fermenters of a-methyl glucoside in segregants from a hybrid yeast. They showed that three distinct genes concerned with the fermentation of α-methyl glucoside were present in the material, and that the rate of fermentation was governed by the presence of these genes either alone or in combination with one another.
Roberts & van der Walt12 recommended that because of the very high frequency of the mutation m → M in Sacch. capensis it should be designated a maltose fermenter. Skinner & Fletcher13 have argued that it seemed reasonable that the difference between no fermentative power and some fermentative power was fundamental, but that qualitative differences between degrees of fermentation were likely to be of trivial importance and to represent only strain differences. Lodder & Kreger-van Rij,8 on the other hand, stated that they did not attach much value to a differentiation between no fermentation and very weak fermentation, while Pappagianis & Phaff11 suggested that the potential ability of yeasts to ferment sugars (such as the slow fermentation of sucrose by Zygosacch. japonicus) should be disregarded for taxonomic purposes and that fermentation should be standardized by using a light inoculum of young cells in order to conceal such ability from detection. It is difficult to follow the reasoning of this last suggestion, as these authors showed that the ability of such yeasts to produce the enzymes necessary for the fermentation of the sugar was a stable characteristic of the yeast, and any stable character of a species should be of taxonomic value. This point of view has been stated convincingly by Barnett2 and by Zsolt,20 who argued that it was inadequate to write + or – for a fermentation, but that a quantitative result should be recorded and that such a result was of value in taxonomy.
It is clear that slow or delayed fermentations can arise from distinct causes, either from adaptation (galactose fermentation in Sacch. chevalieri), mutation (sucrose fermentation in Sacch. capensis), or innately slow fermentation which may be due to very weak production of the required enzyme or to a permeability barrier (sucrose fermentation in Zygosacch. japonicus). Where this ability is a constant and readily demonstrated characteristic of a yeast, then it is of value to record it and, if possible, to determine its cause. One difficulty is to decide how long one should go on looking for these delayed fermentations. Slow galactose fermentation in Sacch. chevalieri manifests itself in 4-6 days as compared to 1-2 days for a normally fermenting Saccharomyces. Brettanomyces will not show fermentation even of glucose until after 3-4 days, and slow maltose fermentation appears in 15-30 days with the Brettanomyces species described here. Kossikov6 found positive fermentation of sucrose by Sacch. globosus appearing after 122 days in sucrose medium. A period such as this is of course impracticable for routine examination of yeasts and it would seem that a reasonable time to allow for the onset of fermentation would be 30 days. It is necessary to distinguish between normal (rapid) fermentations, which should be given primary importance in taxonomy, and the various types of slow or delayed fermentations which should have secondary importance. To do this a quantitative method of measuring the amount of fermentation is required, and the methods available were investigated.
Experimental
Media.—A solution of minerals and vitamins (“MV”) was prepared from: KH3P04, 20 g.; MgSO4, 10 g.; CaC2, 3 g.; pyridoxin, 0-002 g.; thiamine, 0-002 g.; Ca pantothenate, 0-020g.; nicotinic acid 0-002g.; inositol, 0-020g.; biotin, 0-00001 g. and water to 1000 ml. The Base Medium was prepared from: peptone (Bacto), 10 g.; yeast extract (Bacto), 10 g.; “MV” 200 ml. and water to 1000 ml.
Sugar Media were prepared by adding together, under sterile conditions, equal quantities of 8% Seitz-filtered sugar solution and the Base Medium.
Methods for detection of slow fermenters.—
Modifications of or alternatives for standard Einhorn or Durham tube methods for the detection of fermentation have been suggested by Winge & Roberts,19 by Morris & Kirsop,10 by Baird, Hestrin & Lindegren,1 and by Lindegren.7 What is needed is a method which does not miss any fermentations but which does not give false positive results. This method should be simple to operate; it should economize in the use of medium, as some of the rarer sugars are expensive; and it should, if possible, be quantitative. The following methods were selected for testing and were applied to the detection of fermentation in various cases:—
Durham tubes. Test tubes 150 x 18 mm. with inserts 40 X 10 mm. containing 15 ml. of medium.
Durham tubes. Test tubes 100 X 15 mm. with inserts 35 x 8 mm. containing 3 ml. of medium.
Einhorn tubes.
Einhorn tubes sealed with mercury as described by Lindegren.1
Microfermentometer as described by Winge & Roberts.19
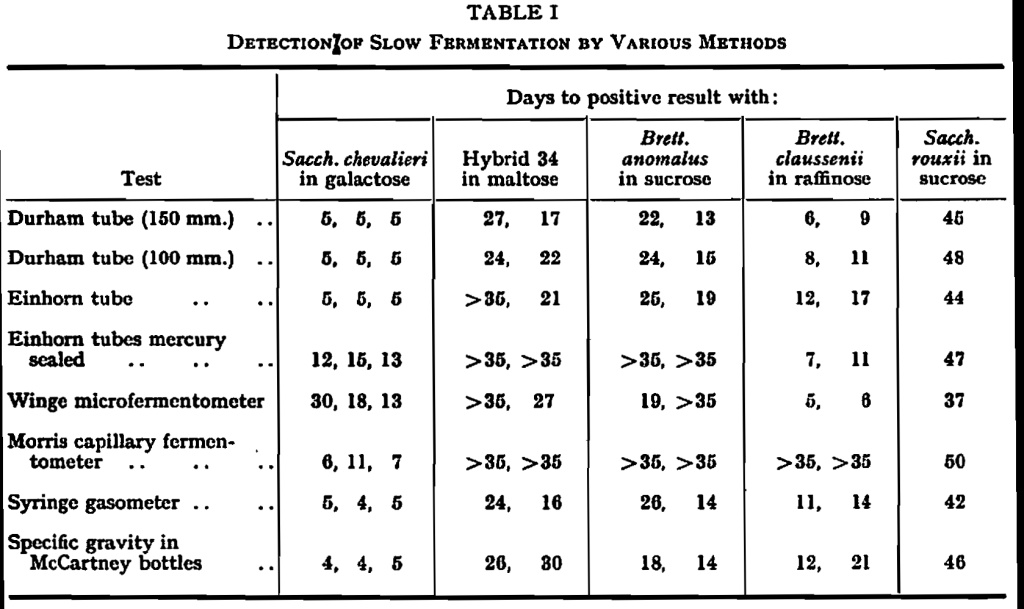
Capillary fermentometer as described by Morris & Kirsop.10
Syringe gasometer as described by Baird, Hestrin & Lindegren.1
25 ml. McCartney bottles containing 3 ml. of the medium for specific gravity determinations at intervals.
From the results of these comparisons, as given in Table I, the following conclusions may be drawn:—
(a) The 150-mm. and 100-mm. Durham tubes were equivalent and were preferable to Einhorn tubes as they gave a positive result more quickly and were also more economical and easier to use. For reasons of economy the 100-mm. test tube was preferred to the 150-mm. test tube.
(b) The mercury-sealed Einhorn was designed to detect constitutive enzymes, and growth in the tube is restricted by the absence of oxygen. In a number of cases this method gave negative results where methods which allowed growth to take place gave positive results, and it was therefore unsuitable.
(c) The micro-methods of Winge or Morris may be preferred for economy of sugar for normal fermentations, but they were not suitable for very slow fermentations, possibly because the anaerobic conditions in these tests prevent yeast growth.
(d) The syringe gasometer gave results equivalent to the Durham tubes, but it had the added attraction that it was quantitative and so allowed very slow continuous fermentations to be distinguished from fast fermentations with a long lag period. For a routine test three bottles were sufficient; these were read daily and lasted till the end of the experiment. Some simplifications of the method were introduced and further tests were carried out with it.
(e) The determination of change of specific gravity during fermentation in McCartney bottles was also quantitative and agreed with the Durham tube results. Where a rapid micro-method for determination of specific gravity was available, such as that described by Williams & Stringer,18 this method was attractive. It had the disadvantage that for protracted tests it was necessary to put on a large number of bottles, as one bottle was sacrificed for each reading.
The syringe gasometer method.—The method of Baird, Hestrin & Lindegren was adapted in the following way:—
The test was carried out in 25-ml. McCartney bottles The aluminium caps of these bottles were specially prepared by drilling in each seven holes, 1 mm. diam., and ensuring that each cap was fitted with a sound rubber liner. 3-ml. quantities of the sugar media were measured into the bottles which were then capped and steam sterilized. With sugars which were known to be unstable under these conditions, 3 ml. of the medium was Seitz-filtered into sterilized bottles which were capped with the special caps under sterile conditions. The bottles were inoculated and incubated at 25°C. Each day the bottle was shaken to release any super-saturation of the medium with gas and the excess gas produced was measured. This was done using a hypodermic syringe, oiled with light mineral oil, and provided with a cut-down No. 18 needle, 15 mm. long. The top of the bottle was swabbed with alcohol and the needle was inserted through one of the holes in the aluminium cap and punctured the rubber seal. If there was any excess pressure in the bottle the syringe piston then moved back. When the piston came to rest it was further moved to the next higher whole ml. reading and this was taken as the amount of gas produced. When the needle was removed the rubber liner sealed itself. A different hole in the cap was used each day of the week so that even in a 35-day experiment one position would not be used more than five times, which the liner was well able to stand without leakage.
A comparison of syringe gasometer results with the fall in specific gravity was made. McCartney bottles containing 3 ml. of sucrose medium were prepared and seeded with Brettanomyces anomalus. The bottles were sampled daily for gas production and, as soon as gas production started, several bottles were used daily for specific gravity determination. The total gas production in individual bottles is compared with the fall in specific gravity in Fig. 1. The oxygen in the gas space in the bottles is presumably absorbed by the yeast for aerobic oxidation of the sugar, with the production of an equal quantity of carbon dioxide. The remainder of the sugar is then fermented releasing 2 molecules of carbon dioxide for each molecule of hexose fermented. This theoretical gas production is included in Fig. 1. The experimental figures fall approximately on a straight line, and it is obvious that it is quite possible to follow the course of a fermentation by this method. The syringe gasometer method is simple and is accurate enough to be of great value for investigating the fermentative ability of yeasts, particularly in the case of very slow fermentations.

Characteristics of the newly isolated yeast.—
The following observations of the characteristics of the new strain were made.
Growth in malt extract. After 5 days at 25° C. cells were of varied shape, round, oval, ellipsoidal or elongated, (2-4) μ X (4-14) μ. Many of the cells had ogive ends. There were many long interlaced mycelia cells, some with true branching “blastese”. The medium was turbid with a fluffy flocculent sediment. A light pellicle with trailing fronds was formed which, on disturbance, fell to the bottom. Growth in malt extract was slow. A characteristic ester smell and flavour developed.
Growth on malt agar. After one month at 15° C. the streak was rough, grained, matter, off-white in colour, with delicate projections at the margin.
Streak culture on malt agar with calcium carbonate. After 2 weeks the streak was rough, grained, off-white. The calcium carbonate was dissolved round the streak. |
Streak culture on glucose-yeast-water-calcium carbonate. The calcium carbonate was dissolved within 10 days.
Slide culture on potato agar. Much pseudomycelium was formed.
Nitrate assimilation. Positive.
Sporulation. No spores were found on gypsum or on Powell’s acetate-agar after one month.
Sugar fermentations. Using the normal Durham tube method with incubation for 10 days the yeast fermented glucose, fructose, mannose, galactose, sucrose, lactose and cellobiose, but not maltose, raffinose, trehalose, melezitose or melibiose.
In extended quantitative tests the following results were obtained.
Maltose was fermented very slowly for 10 days. The rate then increased. Gas was observed in Durham tubes in 14 days. The fermentation was completed in 32 days. On repeated sub-culturing in maltose medium, each subculture being made as soon as gas appeared in a Durham tube, the time to observation of gas production was reduced to 8 days but not further. The yeast therefore cannot adapt to rapid fermentation of maltose.
Trehalose was fermented very slowly at first. After 10 days the rate increased. Gas was observed in Durham tubes after 16 days. The fermentation was completed in 44 days. The yeast could not adapt to rapid fermentation of trehalose as on repeated subculture the time to gas production could only be reduced to 13 days.
Raffinose was fermented extremely slowly at first. After 20 days the rate increased. Gas was observed in Durham tubes after 24 days. The fermentation was completed in 35 days. The yeast could not adapt to rapid fermentation of raffinose as on repeated subculture the time to gas production was not reduced. Only one-third of the raffinose was fermented.
Melezitose and melibiose were not fermented even on extended incubation.
Sugar assimilation. Glucose, fructose, mannose, galactose, sucrose, maltose, lactose, trehalose, raffinose, erythritol, ribose, salicin, cellobiose and α-methyl glucoside were assimilated. Melezitose, melibiose, arabinose, xylose, dulcitol and rhamnose were not assimilated.
Vitamin requirements. Inositol, biotin and thiamine were required. The absence of pantothenate, pyridoxin or nicotinic acid retarded growth.
Characterization of Genus. The small ogive cells, the add production, the slow growth, the production of pseudomycelium, and the characteristic smell and flavor produced in malt extract, were sufficient to characterize this strain as a Brettanomyces.
Comparison with established species.
Differentiation of species in Brettanomyces depends primarily on sugar fermentations. Two species, Brett. bruxellensis and Brett. lambicus were differentiated only by the appearance of their cultures on malt agar, but van der Walt & van Kerken have suggested uniting these two species into a single species, Brett. bruxellensis.
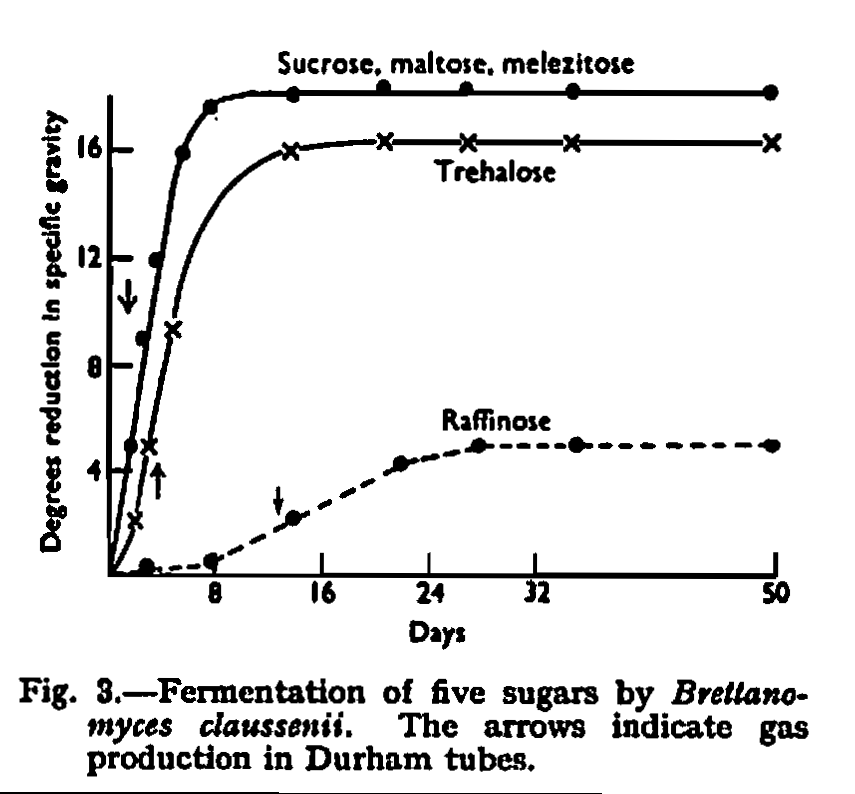
By its ability to ferment lactose the new species was distinguished from all other species except Brett. anomalus and Brett. claussenii which have this property. A detailed comparison was therefore made of the new strain with authentic cultures of Brett. anomalus and Brett. claussenii. These cultures were received from the Centraalbureau voor Schimmel cultures, Delft, and originated from the type cultures studied by Custers. Brett. claussenii agreed in detail with the published description, but Brett. anomalus was found to be completely unable to ferment trehalose; in other respects it agreed with published descriptions. Points of difference shown by this comparison are given in Table II, which does not include characteristics which are common to all three species. The comparative fermentation rates of five sugars are shown in Figs. 2, 3 and 4.

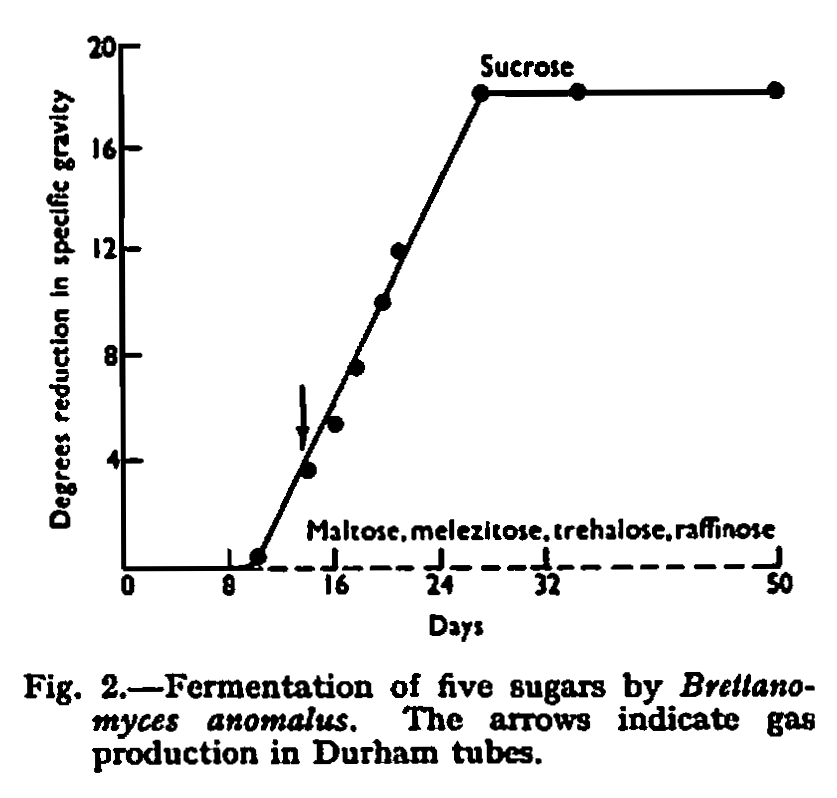
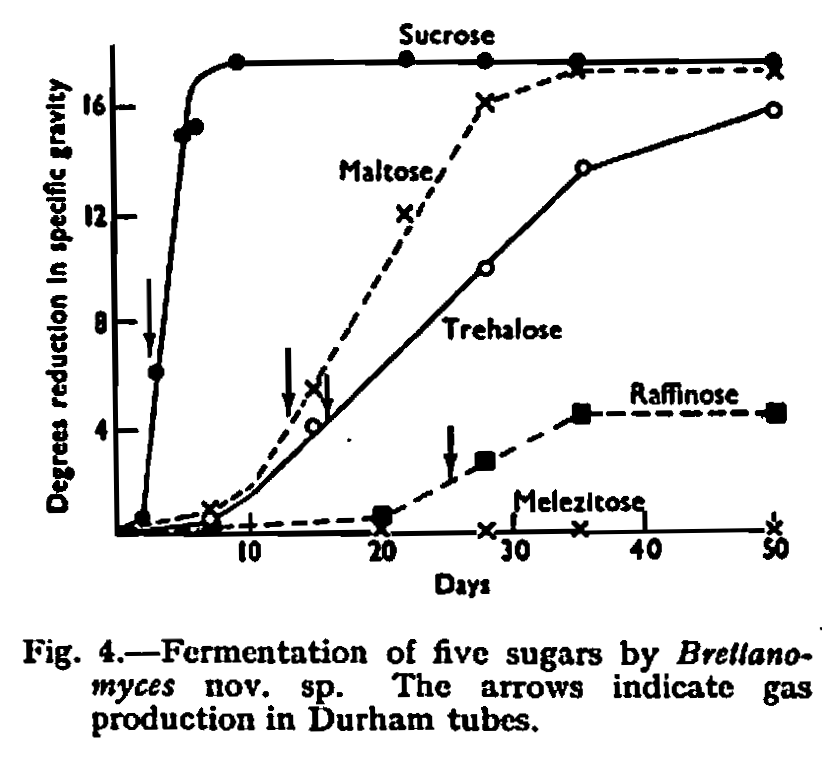
The new strain differed from Brett. anomalus in its ability to ferment sucrose rapidly, in its ability to ferment maltose, raffinose and trehalose slowly, and in its ability to assimilate maltose, erythritol and trehalose.
The new strain differed from Brett. claussenii in its inability to ferment melezitose, in that it could ferment maltose and trehalose only very slowly, in its ability to assimilate erythritol but not melezitose, and in its cultural appearance.
These differences are sufficient to distinguish the strain as a distinct species. The name Brettanomyces dublinensis has been given to the new species and a Latin diagnosis of the species is appended.
Discussion
This investigation has shown the importance of using quantitative fermentation tests for the classification of yeasts. Durham tube tests are ineffective in distinguishing between very slow fermentations and fermentations with a long lag period, or between slow but complete fermentations and weak partial fermentations. Where Durham tubes are used, the period of observation—especially for yeasts in slow-growing genera such as Brettanomyces—should be extended to 30 days. With the yeast under investigation, Durham tube tests terminated at 10 days indicated that the yeast was unable to ferment maltose, trehalose or raffinose. If the tests were extended to 20 days then gas production was observed in maltose and trehalose, but not in raffinose, whereas in 30 days all three of these sugars were fermented.
Some workers have suggested that such slow fermentations should be neglected in taxonomy, but this seems to be unwarranted as the time to observation of fermentation of a sugar was similar each time the test was repeated and on subculture in the same sugar medium the lag period was not eliminated, so these slow fermentations were not due to mutation but were a stable characteristic of the species. On the other hand it would be equally unwarranted to equate the slow fermentations with the fast fermentations and simply call them all positive. The delayed fermentation is a stable distinguishing characteristic, which is readily observed and is therefore of taxonomic value.
In cultural appearance this yeast was rather similar to Brett. anomalus but completely different from Brett. claussenii. In spite of arguments to the contrary it does not seem useful to separate yeast species on morphological appearance alone. In a number of cases both flocculent and non-flocculent strains of a yeast species have been found which are identical biochemically, but which differ in their cultural appearance. For example, flocculent and non-flocculent variants of Saccharomyces cerevisiae and of Sacch. diaslaticus are known, and in the genus Brettanomyces the species Brett. bruxellensis, its variant non-membranaefaciens and Brett. lambicus differ only in cultural appearance, and van der Walt & van Kerken16 have proposed uniting these into a single species. Although it would therefore be wrong to separate the new species on appearance alone from the species Brett. claussenii, nevertheless the difference in appearance is an additional distinguishing characteristic between the new species and Brett. claussenii.
Acknowledgements.—The author wishes to express his thanks to Mr. J. P. Lacey for his technical assistance, to Dr. A. K. Mills for his interest and encouragement, and to the Directors of Arthur Guinness Son & Co. (Dublin), Ltd., for their permission to publish this work.
References
1. Baird. V., Hestrin, S., & Lindegrcn, C. C, Wallerstein Lab. Commun., 1052, IS, 213.
2. Barnctt, J. A., Antonie van Leeuwenhoek, 1957, 23, 1.
3. Gilliland, R. B., this Journal, 1961, 257.
4. Gilliland, R. B., Wallerstein Lab. Commun., 1954,17, 165.
5. Hawthorne, D. C, Heredity, 19C8, 12, 273.
6. Kossikov, K. V., C.I.B.A. Foundation Symposium, Drug Resistance in Micro-organisms,
1957, 102.
7. Lindegren, C. C. Wallcrstcin Lab. Commun., 1950, 19. 40.
8. Loddcr. J.. & Kreger-van Rij. N. J. W., The Yeasts. Amsterdam: North Holland Publishing
Company, 1952.
9. Lodder, J., & Kreger-van Rij. N. J. W., Lab. Practice, 1964, 3. 493.
10. Morris. E. O., & Kirsop. B., this Journal, 1953, 486.
11. Pappagianis, B., & Phaff, H. J., Antonie van Leeuwenhoek, 1050, 22, 353.
12. Roberts. C, & van der Walt, J. P.. C. R. Lab. Carlsberg. 1960. 32, 19.
13. Skinner, C. E.. & Fletcher, D. W., Bad. Rev., 1960. 24, 397.
14. Stelling-Dekker, N. M., Die Sporogenen Hefen, Amsterdam: Verhandel Koninkl. Akad.
Wetenschappen, 1931
15. Takahashi, T., & Ikeda, Y., Z. Vererbungslehre, 1960, 90, 66.
16. van der Walt, J. P., and van Kerken, A. E., Antonie van Leeuwenhoeh, 1950, 25, 146.
17. Wickerham. L. J. V. S., Dept. Agric. Tech. Bull, 1029. 1951.
18. Williams, D.O. & Stringer. W.J. American Brewer, May, 1066, 39.
19. Winge. Ǿ., & Roberts. C. C. R. Lab. Carlsberg, Str. Physiol, 1950, 25, 35.
20. Zsolt, J., Mihrobiologiya, 1060,29 (iii), 278.
