SACCHAROMYCES DIASTATICUS—A STARCH-FERMENTING YEAST
by R. B. Gilliland, Sc.D., F.R.I.C.
(Arthur Guinness, Son & Co. {Dublin} Ltd., St. James’s Gate, Dublin, 8)
Received 29th October. 1966
Work on the yeast tentatively named Saccharomyces diastaticus has justified the separation of starch-fermenting Saccharomyces into a separate species. The name Saccharomyces diastaticus was therefore validated.
Introduction
In 1952 Andrews & Gilliland1 published a description of a yeast which was able to ferment both soluble starch and dextrin and which produced abnormal attenuation of beer. For this yeast the name Saccharomyces diastaticus was proposed. Cultures of this yeast have been distributed widely and it has been studied in many laboratories. Hopkins10-13 investigated the individual enzymes in S. diastaticus and found that its amylolytic property was due to the presence of glucamylase which could promote the hydrolysis of amylose and substrates consisting of unbranched α-(l→4) glucosidic chains. Glucamylase could also break down compounds containing branched chains by by-passing α-(l→6) linkages. This enzyme has also been called amyloglucosidase and it is secreted by Aspergillus niger and Rhizopus delemar but has not been found in any other species of Saccharomyces. Harris, MacWilliam & Phillips9 investigated the attack on the carbohydrates of malt-wort by S. diastaticus. Gilliland6-7 carried out a genetic analysis of this yeast and it has also been used for genetic investigations by Lindegren & Lindegren18, by Windisch & Emeis24, and by Oser.20
In 1943 Bishop & Whitley2 reported the finding of a yeast which secreted diastase; this appears to be the first reported observation of a yeast of the S. diastaticus type. Emeis4 isolated five strains of super-attenuating yeasts from infected beer and one strain from a grape. He commented that yeasts of the type S. diastalicus seemed to be more widely distributed than had hitherto been supposed. Klein, Vacano & Kain16 found that S. diastaticus was the most dangerous and also the most common contaminant of American non-pasteurized bottled beers. They isolated a number of strains and these were examined by Wickerham who found strain differences in agglutination, in ability to attack melezitose, and in tolerance to osmotic pressure. Klein, Vacano & Kain16 also showed that the beer spoilage yeast isolated by Brumsted & Glenister3 originally classified as S. Pastorianus was a variety of S. diastaticus and they suggested that the wild yeast, isolated by Ruff,4 which fermented end-fermented beers was also S. diastalicus. We have isolated S. diastaticus from Irish, English and American beers.
S. diastaticus was different from other species of Saccharomyces in its unique ability to produce and secrete the enzyme glucamylase and this ability was readily determined so that the yeast was easily distinguishable from other species. It was widely distributed and was an important cause of beer spoilage. The original isolates have remained stable in subculture over a period of 20 years. These facts warranted the establishment of S. diastaticus as a separate species. In this paper, some results are quoted on methods of distinguishing between starch fermenting and starch nonfermenting yeasts, justification of species rank for S. diastaticus is discussed.
Emeis4 was unable to confirm differentiation between starch-fermenters and starch non-fermenters on starch-agar plates flooded with iodine, as he observed clear zones round some of the starch non-fermenting yeasts. He confirmed that the yeasts could be unequivocably distinguished by means of their final degree of attenuation in wort or in beer. There have been various conflicting reports of the break-down of starch or dextrins by brewery yeasts. Hopkins showed that in some cases this might be due to the adsorption of amylase by brewer’s yeast from brewery fermentations dressed with malt flour.14 Apart from this, S. cerevisiae contained a debranching enzyme10-13 or isoamylase10,19 which promoted hydrolysis of α-(l→6) linkages in amylopectin and in maltodextrin. This enzyme was not secreted into the medium, so that intact cells of S. cerevisiae had no action on starch or dextrin. On autolysis of the yeast, however, the enzyme was freed and some breakdown of starch might then occur. The action of this enzyme, together with the other yeast enzymes, may account for the clear zones round colonies of starch non-fermenting yeasts on starch-agar plates observed by Emeis.8 There should seldom be any difficulty in distinguishing between the wide zones given by S. diastaticus and the very narrow zones which sometimes appear with S. cerevisiae, but in view of possible confusion the test is not a good one and a more quantitative test is preferable. Emeis4 also failed to demonstrate starch fermentation in a liquid medium consisting of YEP-solution (yeast extract-peptone) with 1% soluble starch in test tubes with Durham inserts. Lindegren & Lindegren18 on the other hand regularly observed fermentation of starch in 5 days and of dextrin in 3 days by the melting-point tube method. In view of the doubts raised by Emeis on the ability of S. diastaticus to ferment starch this was re-examined.
Experimental
Stock cultures of S. diastaticus and S. cerevisiae were sub-cultured into wort in erlenmeyer flasks. They were incubated at 25° C. for 2 days, and the yeast was centrifuged off, washed and then used to inoculate the following media:
Yeast nitrogen base (Difco) + 2% soluble starch
Yeast nitrogen base {Difco) -f- 2% dextrin
These media were contained in 15-ml. amounts in McCartney bottles provided with insert Durham tubes and closed with perforated aluminium caps to enable gas production to be measured by the hypodermic syringe method described by Gilliland.8 The specific gravities of the media were determined at the beginning of the experiment and at intervals thereafter by the falling drop method of Williams & Stringer.25
In view of the failure of Emeis to detect starch fermentation in 1% starch medium by the standard Durham tube method, this method was also tested using 1 and 2% soluble starch medium in cotton wool plugged test tubes with Durham tube inserts.
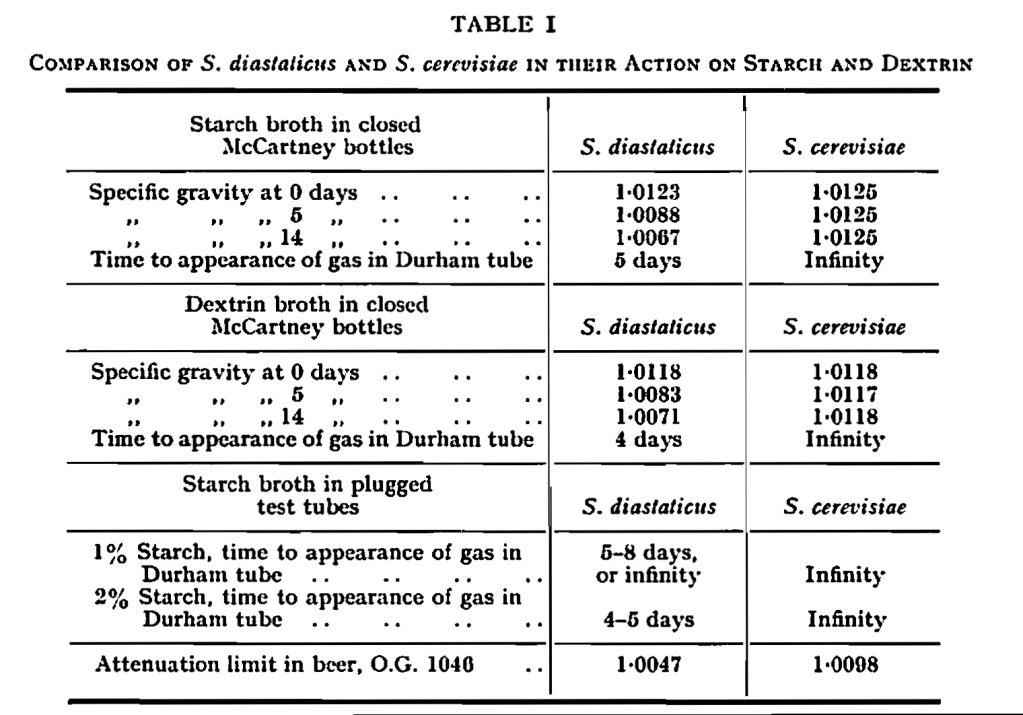
The course of fermentations as demonstrated by gas production in McCartney bottles is shown in Fig. 1. It was clear that the fermentations were slow and continued over about 14 days. Gas was observed in insert Durham tubes after 5 days in starch and after 4 days in dextrin. In the same time the specific gravities of the media had fallen significantly as is shown in Table I. No gas was produced, and there was no fall in specific gravity, when these media were seeded with S. cerevisiae and incubated under similar conditions.
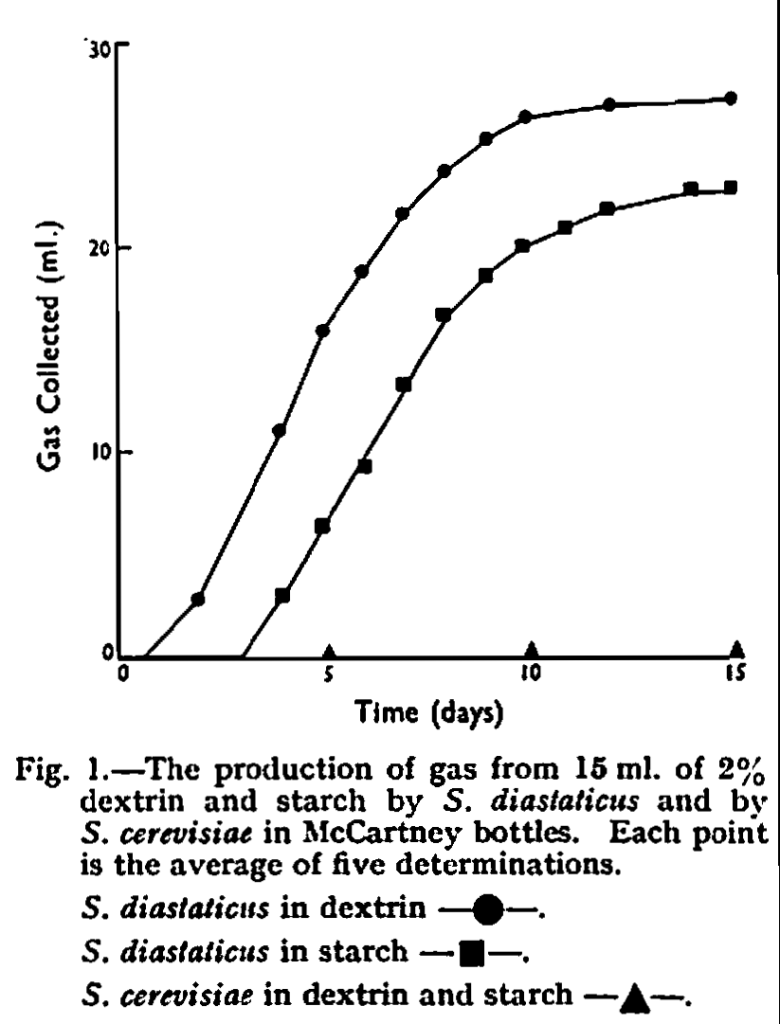
Gas was observed in the standard Durham tube method in 2% soluble starch seeded with S. diastaticus in 4-5 days and the insert tube was two-thirds full in 7 days. With 1% starch, on the other hand, observation of gas was irregular and in a set of similar tubes some showed gas in 5-8 days while occasional tubes never showed gas. If the seeding rate was low, gas was not observed. The problem of the detection of slow fermentations was fully discussed in an earlier paper8 in which it was shown that negative results were readily obtained in the standard Durham tube tests if gas was produced slowly. In this case the gas diffused to the open surface of the liquid and escaped there so that no bubbles were collected in the insert tube. It was therefore essential that a quantitative test such as the hypodermic syringe method, or the specific gravity method, should be used for slow fermentations.
S. diastaticus could not attack raw unheated starch with unbroken grains. If the medium containing the starch was sterilized then the starch became fermentable. This is readily understandable as amylase will not normally attack unbroken starch.
Discussion
The experimental results demonstrated that soluble starch and dextrin were fermented by S. diastaticus but not by S. cerevisiae. They also confirmed that super-attenuation in beer was dependent on the ability to ferment dextrin, as indeed it must be, and that in S. diastaticus this was associated with ability to ferment starch. Difficulties reported in detecting starch fermentation by S. diastaticus may have been due to the inefficiency of the standard Durham tube method in detecting slow fermenters particularly at low carbohydrate content in the medium.
Apart from starch fermentation the other distinguishing characters of S. diastaticus were its resistance to high temperatures, its cell shape, and its variability in cell size. The question remained whether these were sufficient characteristics to distinguish it as a species. Species in the genus Saccharomyces have traditionally been distinguished mainly on their ability to ferment glucose, sucrose, galactose, maltose, raffinose, melibiose, and lactose. This selection of sugars is rather arbitrary and additions to the list have been suggested by various authors. If a wide variety of sugars was used it might be possible to split many species of Saccharomyces but this would clearly be undesirable, especially if it entailed the use of either rare sugars or sugars with little biological significance. On the other hand the ability or lack of ability to ferment starch, which is one of the commonest of all carbohydrates, is of great importance both biologically and technologically and it should rate very high as a distinguishing characteristic.
Kudriavzev17 has attempted to produce an evolutionary pattern for species in the genus Saccharomyces and in his diagram of the phylogenesis of these yeasts he left a space for a yeast with the ability to ferment complex dextrins. S. diastaticus fits perfectly into this space. So whether on simple technological usefulness, or normal distinguishing tests for a species, or on phylogenetic principles, a strong case can be made for species rank for S. diastaticus. Variations within the species have been noted. These include ability to flocculate,1,5,16 ability to attack melezitose,4 and tolerance to osmotic pressure.16 Such variations do not in any way invalidate the species. If they were considered of sufficient importance the species might be split but at present there seems no justification for this.
To validate the species, van der Walt23 published an abbreviated Latin diagnosis but fuller diagnoses are provided below.
Saccharomyces diastaticus
After 3 days in malt extract at 25° C. the cells were round to oval (3-7) μ X (4-10) μ, single or in pairs; a sediment was formed; no pellicle was observed, but after 1 month a slight ring was present. After 3 days on malt agar at 25° C. cells were round to oval (3-7) μ X (4-10) μ single or in pairs; after 1 month at 17° C. the streak was cream coloured, soft, smooth, and shining. No pseudomycelium was formed on potato agar. The yeast spored readily on gypsum blocks, the spores were round, 1-4 per ascus, size 3 μ x 3 μ; no conjugation occurred prior to ascus formation. Glucose, galactose, sucrose, mannose, maltose, raffinose (one-third), dextrin and starch were fermented. Lactose, trehalose, cellobiose, α-methyl glucoside, xylose, arabinose and melezitose were not fermented. Glucose, galactose, sucrose, mannose, maltose, raffinose, dextrin, soluble starch, trehalose and melezitose were assimilated in a synthetic medium. Lactose, cellobiose, a-methyl glucoside, xylose and arabinose were not assimilated. Potassium nitrate was not assimilated. Good growth was obtained with alcohol as the sole source of carbon. Biotin, pantothenate and inositol were required for growth in synthetic media. Growth was obtained at 37° C. The yeast was isolated from beer.
Variants within the species may be strongly agglutinated, or may ferment melezitose, or raffinose completely instead of one-third.
References
I. Andrews. J. & Gilliland. R. B., this Journal, 1052, 180.
2. Bishop, L. R., & Whitloy, N. A., this Journal. 1043, 223.
3. Brumsted, D. D, & Glenistcr, P. R., Proc. A. M. Amer. Soc. Brew. Chem., 1962, 72.
4. Emeis. C, Monats. Brau., 1063, 16, 33.
5. Gilliland, R. B., Walterstein Lab. Commun., 1064, 17, 166.
6. Gilliland, R. B., Nature, Lond., 1064.173, 400.
7. Gilliland, R. B., Proc. Eur. Brew. Conv. Nice. 1963, 121.
8. Gilliland, R. B., this Journal, 1062, 61.
9. Harris, G. MacWilliam, I. C, & Phillips, A. W., Proc. Eur. Brew. Conv. Copenhagen, 1057.
173.
10. Hopkins, R. H., Proc. Eur. Brew. Conv. Baden-Baden, 1966, 62.
11. Hopkins, R. H.. & Kulka. D., Proc. Eur. Brew. Conv. Copenhagen, 1067, 182.
12. Hopkins, R. H., & Kulka. D., Arch. Biochem. Biophys., 1967, 69, 46.
13. Hopkins, R. H., Wallerstein Lab. Commun., 1068, 21, 300.
14. Hopkins, R. H., this Journal, 1066. 427.
15. Kerr, R. W.. Cleveland, F. C, & KaUbeck, J., J. Amer. chem. Soc., 1951,73, 3916.
16. Klein, J. G. Vacano, N. L. & Kain, N. A., Proc. A. M. Amer. Soc. Brew. Chem., 1964,
166.
17. Kudriavzev. V. I., Yeast Systematics. Academy of Moscow, 1064.
18. Lindegren, C. C, & Lindegren, G., J. gen. Microbiol., 1066,15, 19.
19. Manners, D. J., & Khin Maung., Chem. & Ind., 1955. 050.
20. Oser, H., Archiv. für Mikrobiologie, 1082, 44, 47.
21. Phillips, L. L., & Caldwell, M. L., /. Amer. chem. Soc., 1061,73, 3550 and 3663.
22. Ruff, D. G.. Brewers’ Digest, 1068, 33, 62.
23. van der Walt, J. P., Antonie van Leeuwenhoek, 1065,31,187.
24. Windisch, S., & Emeis, C. C, Brauerei, Wissensch. Beil.. I960, 13, 2.
25. Williams, D. O., & Stringer, W. J.. Amer. Brewer, May, 1965, 39.
